A15 - Funktionen von endozytotischen c-Cbl- und Hakai-Proteingerüsten im vesikulären Transport von Rezeptoren der Zelloberfläche
Projektleiter
Prof. Dr. Walter Birchmeier, MDC
Prof. Dr. Matthias Selbach, MDC
Ein entscheidender Prozess in der Embryonalentwicklung sowie auch bei der Metastasierung von Tumoren ist die epitheliale-mesenchymalen Transition (EMT), ein zellulärer Prozess bei welchem epitheliale Zellen ihre Morphologie massiv verändern und Fibroblasten-ähnliche Eigenschaften annehmen. Der Verlust der Zelladhesion wird dabei über die Expressionsdynamik von E-Cadherin und weiteren assoziierten Proteine reguliert. E-cadherin spielt als Adhesionsmolekül eine wichtige Rolle bei Stabilisierung von Zell-Zell-Kontakten, Erhaltung der Zellpolarität, als auch bei der Progression von Tumoren. Die Expressionsdynamik von E-Cadherin kann durch Stimulierung von Rezeptor-Tyrosinkinase Signalwegen beeinflusst werden und zu einer EMT sowie erhöhter Zellmotilität führen1-6.
Im Labor von Prof. W. Birchmeier wurde das Protein Hakai entdeckt und es konnte gezeigt werden, dass Hakai mit tyrosinphosphoryliertem E-cadherin interagiert und dessen Ubiqitinierung und die Endozytose des E-cadherin/β-catein Membran Komplexes induziert 7. Die Expression von Hakai in Epithelzellkulturmodel stört die Zell-Zell Adhesion, verstärkt die Endozytose von E-Cadherin und steigert die Zellmotilität. Demnach spielt Hakai eine wichtige regulatorische Rolle in der epithelialen-mesenchymalen Transition während Embryonalentwicklung als auch Metastasierung7. Hakai ist sowohl strukturell als auch funktionell verwandt mit c-Cbl, ebenfalls ein membranassoziiertes Multidoman-Scaffolding-Protein mit E3 Ubiquitin Ligase Aktivität. Hakai und c-Cbl sind an Prozessen der Rezeptor-Tyrosinkinase induzierten EMT beteiligt und regulieren die Endozytose sowie intrazellulären Vesikeltransport von Zelladhesionsmolekülen und Zelloberflächenrezeptoren (Fig. 1).
Die c-Cbl und Hakai-abhängige Regulation räumlicher und zeitlicher Dynamik von Zellmembranproteinen (z.B. E-Cadherin) ist bisher nicht komplett aufgeklärt. Zudem ist die molekulare Basis für die nicht-redundanten zellulären Funktionen der beiden strukturell so ähnlichen Proteine nicht bekannt.
Die im Labor von Prof. M. Selbach bereits etablierte Methode der quantitative Proteomanalyse mittels Massenspektrometrie bietet einen viel versprechenden Ansatz für die Untersuchung komplexer biologischer Prozesse, im besonderen bei der Identifizierung von Protein-Protein Interaktionen 89,10.
Ziel diese Forschungsprojektes ist es, mittels genetischer, molekularbiologischer und biochemischer Ansätze, die gemeinsamen und unterschiedlichen Funktionen von c-Cbl und Hakai aufzuklären. Der kombinierte Einsatz von quantitativer Proteomics (Fig. 2), Mikroskopie und in vivo-Modellen wird einen direkten Verlgeich der posttranslationalen Modifikationen, Interaktionspartnern, sowie von Phenotypen entsprechender Knock-out-Mäuse ermöglichen. Die Unterschiede und Übereinstimmung der molekularen Mechanismen und Funktionen von Hakai und c-Cbl sowie deren Integrität in der Pathogenese humaner Erkrankungen werden dabei aufgeklärt.
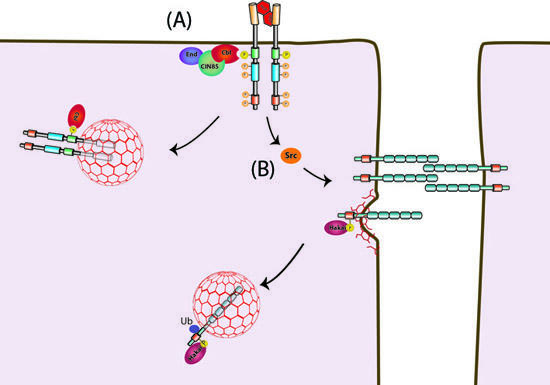
Hakai and Cbl regulate endocytosis
Hakai and Cbl are recruited to phosphorylated cell-membrane proteins, such as E-cadherin or Met, and trigger their internalization. (A) Activation of the receptor-tyrosine-kinase-receptor Met results in activation of the kinase Src and in recruitment of Cbl. While Cbl regulates the internalization of Met, (B) Src phosphorylates the cell adhesion molecule E-cadherin and generates a binding site for Hakai. Recruited Hakai induces ubiquitination and endocytosis of E-cadherin.
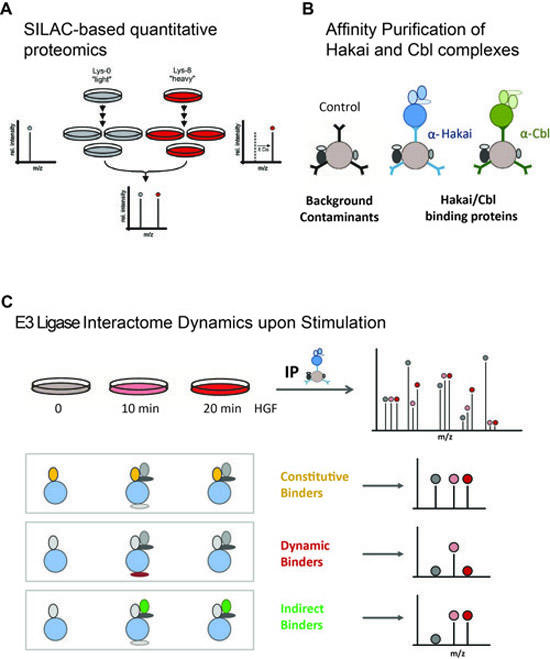
Experimental Approach to compare Hakai and Cbl Interactomes.
SILAC labeling of mammalian cell culture (A) allows for precise and accurate quantification of protein abundance. Using this technology, Hakai and Cbl protein complexes (B) will be studied by quantitative affinity purification followed by mass spectrometry (q-AP-MS).
In addition, dynamic changes of these two E3 ligase interactomes upon cell activation will be investigated by capturing protein complex composition at various time points upon growth factor (HGF) stimulation (C).
References:
- Perl, A.K., Wilgenbus, P., Dahl, U., Semb, H. & Christofori, G. A causal role for E-cadherin in the transition from adenoma to carcinoma. Nature 392, 190-3 (1998).
- Behrens, J., Mareel, M.M., Van Roy, F.M. & Birchmeier, W. Dissecting tumor cell invasion: epithelial cells acquire invasive properties after the loss of uvomorulin-mediated cell-cell adhesion. J Cell Biol 108, 2435-47 (1989).
- Heuberger, J. & Birchmeier, W. Interplay of cadherin-mediated cell adhesion and canonical wnt signaling. Cold Spring Harb Protoc 2, a002915 (2010).
- Behrens, J. et al. Loss of epithelial differentiation and gain of invasiveness correlates with tyrosine phosphorylation of the E-cadherin/beta-catenin complex in cells transformed with a temperature-sensitive v-SRC gene. J Cell Biol 120, 757-66 (1993).
- Weidner, K.M., Behrens, J., Vandekerckhove, J. & Birchmeier, W. Scatter factor: molecular characteristics and effect on the invasiveness of epithelial cells. J Cell Biol 111, 2097-108 (1990).
- Riethmacher, D., Brinkmann, V. & Birchmeier, C. A targeted mutation in the mouse E-cadherin gene results in defective preimplantation development. Proc Natl Acad Sci U S A 92, 855-9 (1995).
- Fujita, Y. et al. Hakai, a c-Cbl-like protein, ubiquitinates and induces endocytosis of the E-cadherin complex. Nat Cell Biol 4, 222-31 (2002).
- Vermeulen, M. & Selbach, M. Quantitative proteomics: a tool to assess cell differentiation. Curr Opin Cell Biol 21, 761-6 (2009).
- Selbach, M. et al. Host cell interactome of tyrosine-phosphorylated bacterial proteins. Cell Host Microbe 5, 397-403 (2009).
- Selbach, M. & Mann, M. Protein interaction screening by quantitative immunoprecipitation combined with knockdown (QUICK). Nat Methods 3, 981-3 (2006).
